Il Metabolismo del Solfuro di Idrogeno nel cancro
Le cellule tumorali sono strettamente dipendenti dalla glicolsi aerobica (effetto Warburg) e dall’amminoacido metionina (Effetto Hoffman). L’effetto Warburg viene chiamata glicolisi aerobica, perché utilizzano il glucosio e producono acido lattico anche in presenza di ossigeno. La presenza o meno dell’ossigeno, sembra non inteferire con il metabolismo glicolitico del tessuto tumorale; come mai? A questa domanda risposi in diverse pubblicazioni precedenti: I mitocondri delle cellule tumorali sembrano che non respirano l’ossigeno, ma respirano l’acido fumarico.
Gli articoli dedicati a questo argomento sono i seguenti:
- La Respirazione del Fumarato nei tumori https://liborioquinto.altervista.org/la-respirazione-del-fumarato-nei-tumori/
- Un possibile meccanismo dell’effetto Warburg https://liborioquinto.altervista.org/un-possibile-meccanismo-delleffetto-warburg/
- H2S: Tra longevità e cancro https://liborioquinto.altervista.org/h2s-tra-longevita-e-cancro/
- Il Ruolo della Cisteina diossigenasi nel cancro https://liborioquinto.altervista.org/il-ruolo-della-cisteina-diossigenasi-nel-cancro/
Le cellule tumorali sono anche fortemente dipendenti dalla metionina, e questa dipendenza prende il nome di effetto Hoffman; Gli studi su linee cellulari di cancro e modelli animali hanno dimostrato che le diete a forte restrizione di metionina “associate ad iniezioni nel sangue di enzima METIONINASI”, sembrano essere in grado di inibire la proliferazione cellulare. Mentre le cellule sane sono in grado di riciclare l’omocisteina e la Metiltioadenosina (MTA) in metionina, le celluletumorali non sono capaci, e quindi per vivere consumano grandi quantità di metionina. Le cellule tumoarali sono carenti di METIONINA SINTASI e questa carenza impedisce alle cellule tumorali di riciclare l’omocisteina in metiionina; le cellue tumorali sono anche carenti dell’enzima S-Metil-5-tioadenosina-fosforilasi (EC2.4.2.28) e quindi non possono riciclare la MTA in Metionina; la carenza di quetsi due enzimi rende la cellula tumorale fortemente dipendente dalla metionina dietetica e quindi la sua restrizione potrebbe inibire la crescita dei tumori.
Sulla dipendenza dei tumori dalla metionina ho dedicato vari articoli, che invito il lettore a leggere:
- Perché le cellule tumorali sono avide di metionina? https://liborioquinto.altervista.org/perche-le-cellule-tumorali-sono-avide-di-metionina/
- Effetto Hoffman nel cancro https://liborioquinto.altervista.org/effetto-hoffman-nel-cancro/
- Dipendenza del cancro dalla metionina dietetica https://liborioquinto.altervista.org/dipendenza-del-cancro-dalla-metionina-dietetica/
I Tumori hanno un grande bisogno di metionina principalmente per due motivi:
- I tumori utilizzano la metionina per produrre METILTIOADENOSINA (MTA) che è un potente soppressore del sistema immunitario innato, ossia addormenta i MACROFAGI. Per essere più tecnici si dice che i tumori producono MTA dalla metionina per trasformare i Macrofagi antitumorali M1, in Macrofagi protettivi del tumore M2. In poche parole i tumori corrompono i Macrofagi attraverso la produzione di MTA. I Tumori addormentano il sistema immunitario anche con la CHINURENINA un metabolita del TRITOFANO; in questo caso la CHINURENINA va ad addormentare i NATURAL KILLER e i linfociti-T citotossici.
- I Tumori utilizzano la metionina per produrre solfuro di Idrogeno, un gas che consente alle cellule tumorali di riprogrammare il metabolismo mitocondriale, passando dalla respirazione aerobica alla respirazione del fumarato. Quando la cellula tumorale incomincia a respirare il fumarato al posto dell’ossigeno, si attiva l’effetto Warburg: La cellula inizia a consumare glucosio per convogliarlo nella glicolisi e convertirlo in Ossalacetato attraverso la PEPC (Fosfoenolpiruvato carbossilasi). L’ossalacetato attraverso il ciclo di Krebs inverso andrà a formare acido fumarico che verrà respirato dal mitocondrio al posto dell’ossigeno. Cosa significa respirare? In genere le cellule respirano l’ossigeno, ossia utilizzano l’ossigeno come accettore terminale di elettroni, producendo acqua; Nel caso dei tumori, i mitocondri utilizzano l’acido fumarico come accettore di elettroni, producendo come sottoprodotto l’acido succinico. L’acido succinico è infatti uno degli oncometaboliti più abbondanti nel microambiente tumorale dopo l’acido lattico.
La forte dipendenza delle cellule tumorali dal glucosio ha come principale scopo quello di produrre acido fumarico necessario ai mitocondri tumorali per respirarlo al posto dell’ossigeno.
Quindi ripetiamo:
Le cellule tumorali consumano grandi quantità di metionina per produrre solfuro di idrogeno; il solfuro di idrogeno blocca il complesso-4 della catena di trasporto degli elettroni, ed il flusso degli elettroni viene dirottato verso la SUCCINATO DEIDORGENASI (COMPLESSO-2) che dventa FUMARATO REDUTTASI. La fumarato reduttasi scarica gli elettroni dell’ETC direttamente sull’acido fumarico, producendo acido succinico.
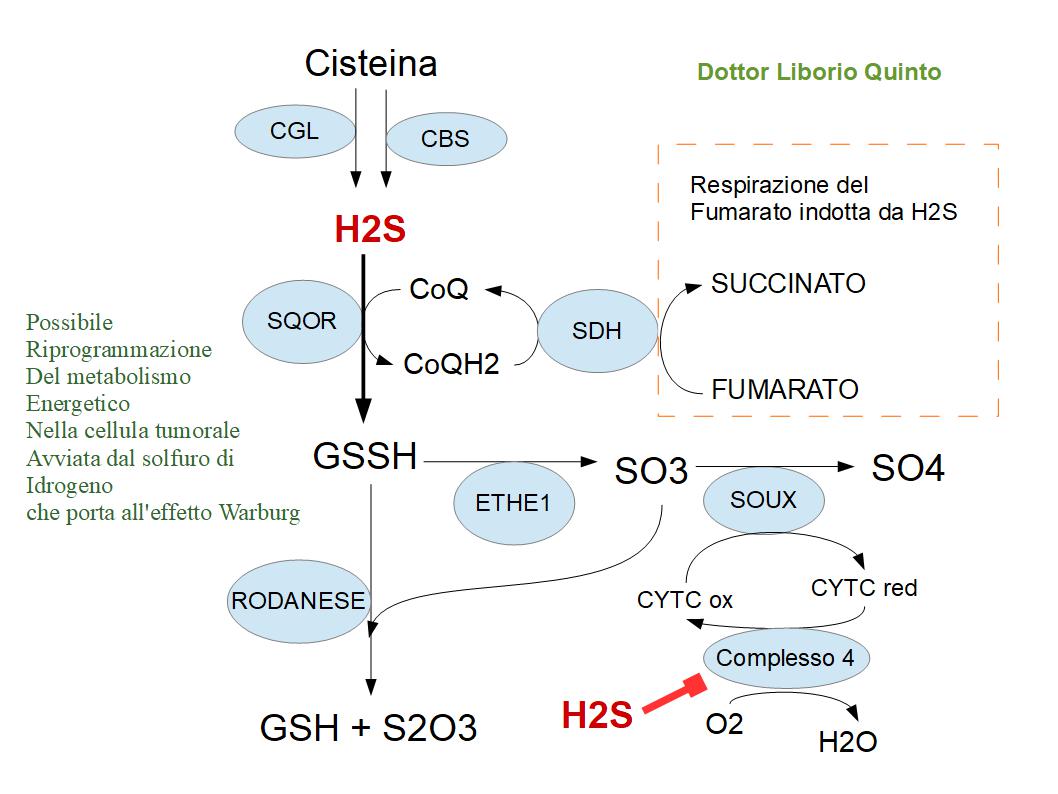
Il Metabolismo del Solfuro di Idrogeno nel cancro
Il solfuro di idrogeno viene prodotto dagli enzimi CISTATIONINA GAMMA-LIASI (CGL) e CISTATIONINA BETA-SINTASI (CBS).
Il solfuro di idrogeno a certi dosaggi inibisce il complesso-4 dell’ETC, ossia il citocromo-c-ossidasi; di conseguenza i mitocondri delle cellule tumorali perdono la capacità di utilizzare l’ossigeno come accettore di elettroni e ciò spiega l’effetto Warburg delle cellule tumorali “chiamata anche glicolisi aerobica“, dove la glicolisi con produzione di lattato, avviene anche in presenza di ossigeno.
Il solfuro di idrogeno viene anche sfruttato dalla cellula tumorale per produrre energia ATP; infatti sulla membrana interna mitocondriale è presente l’enzima SQOR che ossida il solfuro di idrogeno per ridurre il CoQ10 (ossidato) in CoQ10 (ridotto); Il CoQ10 (ridotto) verrà riciclato in CoQ10 (ossidato) dalla FUMARATO REDUTTASI, che scaricherà gli elettroni direttamente sul FUMARATO (respirazione del fumarato).
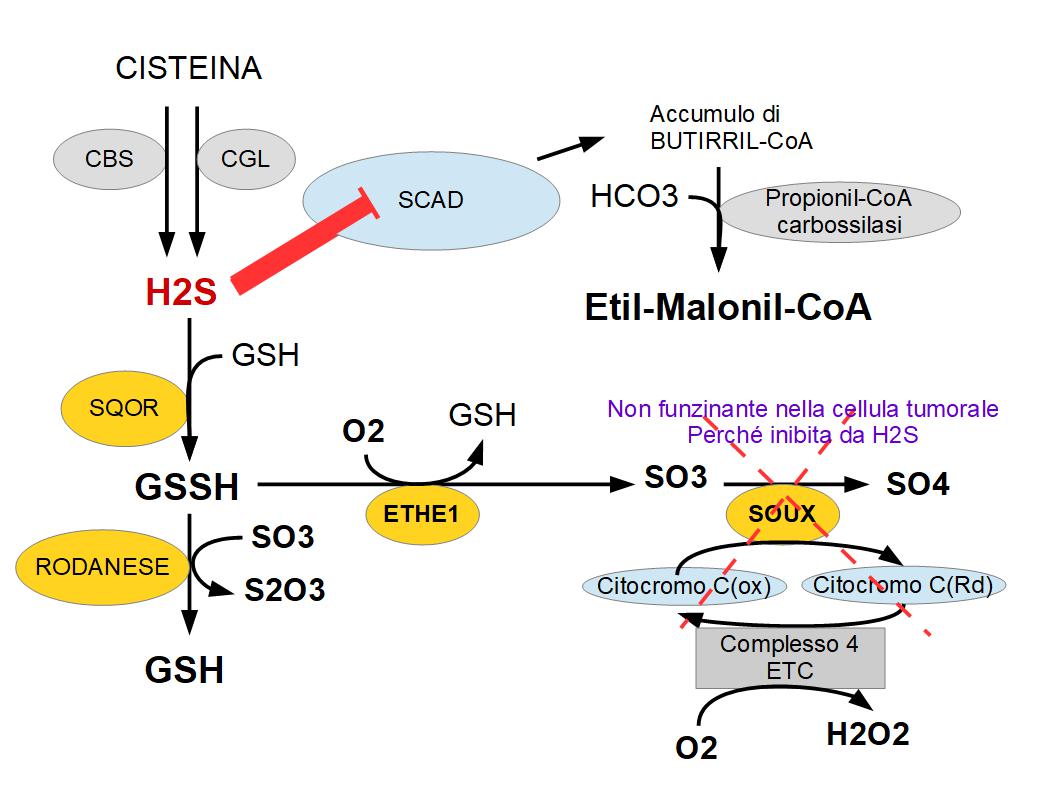
Il sottoprodotto dell’enzima SQOR è il GLUTATIONE SULFINATO “GSSH”, che normalmente viene poi trasformato dall’enzima ETHE1 in solfito e dalla SOUX (solfito ossidasi) in solfati. Nelle cellule tumorali questo percorso “ETHE-1 – SOUX” è completamente bloccato, perché SOUX non può trasformare i solfiti in solfati se il Complesso-4 è fuoriuso; Infatti abbiamo detto che nei tumori, il solfuro di idrogeno sopprime il complesso-4 della ETC. SOUX infatti lavora insieme al Citocoromo-C. SOUX ossida i solfito in solfato; e contemporaneamente riduce il citocromo-C, il quale viene riossidato cedendo gli elettroni al complesso-4 che a sua volta scarica gli elettroni sull’ossigeno, producendo acqua; ma nei tumori siccome il complesso-4 è soppresso dal solfuro di idrogeno, SOUX non può funzionare; Ne consegue che il solfito viene ossidato in TIOCIANATO da un altro enzima : IL RODANESE. Il Rodanese normalmente ha la funzione di detossificare le cellule dal cianuro: Utilizza il tiosolfato S2O3-) per trasformare il cianuro HCN- in CNS- (tiocianato) + Solfito; Ma nelle cellule tumorali poiché ETHE1 produce una grande quantità di solfito non ossidato da SOUX, la RODANESE tumorale è obbligata a catalizzare la reazione inversa, ossidando il GLUTATIONE SULFINATO GSSH e il solfito in glutatione GSH + Tiosolfato (S2O3).
Il Solfuro di Idrogeno nelle cellule tumorali, provoca anche la produzione di Acido etil-malonico; Il solfuro di idrogeno infatti inibisce l’enzima SCAD (EC1.3.99.2) chiamato anche acil-coA deidrogenasi a catena corta; quetsa inibizione impedisce l’ossidazione del BUTIRRIL-CoA, che verrà metabolizzato dagli enzimi PROPIONIL-CoA carbossilasi e/o dalla ACETIL-CoA carbossilasi in ETIL-MALONIL-CoA, provocando aciduria etil-malonica nel microambiente tumorale. ETIL-MALONIL-CoA è un inibitore di molti enzimi della catena di trasporto degli elettroni.
Un’altra propietà antitumorale della Vitamina C
Abbiamo detto che il solfuro di idrogeno inibisce il complesso-4 della catena di trasporto degli elettroni; Se non funziona il complesso-4, non può funzionare neanche la SOLFITO OSSIDASI (SOUX), e quindi i solfitiprodotti da ETHE1 vengono dirottati verso la RODANESE. La Vitamina C pare attenuare l’inibizione del compelsso-4 da parte del solfuro di idrogeno. Infatti alla pubblicazione scientifica PMC3579397 viene affermato che la somministrazione di DHA “la forma ossidata della vitamina C” nei mitocondri di ratto previene l’inibizione del complesso-4 da parte del solfuro di idrogeno.
Acido Etil-Malonico nel cancro protrebbe ridurre i livelli di fosfocreatina.
Abbiamo detto che nei tumori i solfiti prodotti da ETHE1 non possono essere ossidati dalla SOUX perché il complesso-4 è bloccato dal solfuro di idrogeno; SOUX può ossidare il solfito in solfato se e soltanto se il complesso-4 può utilizzare l’ossigeno come accettore di elettroni, ma nei tumori non è possibile per colpa del solfuro di idorgeno. Ne consegue che nel mitocondrio tumorale si accumulano solfiti e SOLFURO DI IDROGENO. L’eccesso di solfuro di idrogeno non suficientemente smaltito dalla cellula tumorale, va ad inibire l’acil-CoA deidrogenasi a catena corta “SCAD (EC1.3.99.2)”; ciò determina un accumulo di BUTIRRIL-CoA che verrà trasformato in maniera anomala in ACIDO ETIL-MALONICO dai enzimi Acetil-CoA carbossilasi e Propionil-CoA carbossilasi”.
L’acido ETILMALONICO a sua volta potrebbe andare ad inibire la FOSFOCREATINA CHINASI CPK, riducendo i livelli di ATP a pronto utilizzo. (Dati Brenda Enzyme EC1.3.99.2)
Fonte Bibliografica
- Tiranti V, Zeviani M. Altered sulfide (H(2)S) metabolism in ethylmalonic encephalopathy. Cold Spring Harb Perspect Biol. 2013 Jan 1;5(1):a011437. doi: 10.1101/cshperspect.a011437. PMID: 23284046; PMCID: PMC3579397.